ゾウムシ上科の系統分類学概説
2008年6月5日 小島 弘昭
ゾウムシ類は,頭部の一部が伸長し,その先に口器を備えユニークな形をした甲虫で,今から約1億5千万年前のジュラ紀に,この口吻を獲得し,姉妹群であるハムシ上科から分化し起源した.ゾウムシは,その後,口吻に“産卵管”的な機能を加えるとともに,地球上で繁栄した被子植物を利用することで適応放散し,これまでに約5,700属60,000種(推定種数20万種)が知られる生物界でも最大級の分類群にまで多様化した.
ゾウムシの魅力は,その特徴である口吻に秘められているといっても過言ではなく,「他の虫より頭が前に伸びた分だけ面白みが詰まっている」,とはゾウムシ研究の大家,森本桂博士の格言である.チョウ類やカメムシ類における吸汁式口器の獲得や,ハチ類に見られる産卵管形態の進化による繁栄と同様,ゾウムシ類における口吻の獲得は,この類の繁栄を導いた.
ゾウムシ上科の分類
ゾウムシ上科の分類体系については,いまだ統一した見解が得られていない.最近では,キクイムシ科,ナガキクイムシ科をゾウムシ科に含める研究者が多い.同じような生息環境にいて,外見がよく似たキクイゾウムシ亜科から派生したグループとして扱われるが,キクイゾウムシ+ゾウムシとキクイムシ,ナガキクイムシでの頭部構造の根本的違いを認識しておらず,独立の科として扱うのが妥当であろう.科の分類は,7つとする研究者から,20とする研究者まで様々である.
ゾウムシ上科各科(あるいは各科群)の概説
チョッキリモドキ科(Nemonychidae):成虫,幼虫ともに,おもにナンヨウスギ,マツ,イヌマキなど針葉樹の花粉食で,北南両半球の温帯域に隔離分布する遺存的な分布パターンを示す.現生種は3亜科(Nemonychinae, Rhinorhynchinae, Cimberidinae)からなり,70種程度が知られる.旧北区東部からは知られていないが,将来,日本からも発見される可能性がある.化石は上部ジュラ紀から知られている.
ヒゲナガゾウムシ科(Anthribidae):成虫は基本的に菌食性で,一部,種子食,捕食者も知られる.発達した産卵管を有し,倒木や朽ち木,キノコに産卵する際,口吻で傷を付け,あとは産卵管を使って卵を産み込む.3亜科(Anthribinae, Choraginae, Urodontinae)約3,000種からなり,熱帯域を中心に分布する.最も古い化石は白亜紀中期から知られる.
アケボノゾウムシ科(Belidae):寄主植物,食性は多様で,ソテツ,針葉樹,ヤシなどの生殖器官を利用するものや,シダ植物から得られるものもある.オーストラリアでは,アカシアやユーカリなど被子植物を利用するものも多いが,形態的にも祖先的なグループは,針葉樹を利用し,幼虫が枯れ枝や朽ち木内に生息する.現生種は3亜科(Belinae, Oxycoryninae, Aglycyderinae)約400種が知られ,南半球を中心に分布し,一部が熱帯域からも知られる.研究者によっては,各亜科を科レベルに昇格して扱うこともある.上部ジュラ紀から知られるEobelinaeは本科の化石群とされている.
ミナミホソクチゾウムシ科(Caridae):現生種はオーストラリアと南米南部からのみ知られ,ヒノキ科植物を利用し,幼虫は雌の球果内で種子を食べることが知られている.既知種は10種に満たない.上部ジュラ紀から知られるEccoptarthinaeは,本科の化石群と考えられている.
オトシブミ科(Attelabidae):植物の茎を切断,葉を切って揺籃をつくるなど,口吻を用いて産卵時に何らかの加工を行う.オトシブミの一部には,胞子の貯蔵器官を腹部に有し,産卵時,揺籃に植えつけることが知られており,この仲間の幼虫は,健全な植物組織を利用せず,食性は腐食か菌食性と考えられる.おもに被子植物を利用するが,チョッキリゾウムシの仲間にマツやヒノキなど針葉樹を利用するものも少ないながら含まれる.2亜科(Attelabinae, Rhynchitinae)約2,000種が熱帯から温帯にいたる地域に分布するが,ニュージーランドやニューカレドニア,ハワイなど大陸から離れた島には分布しない.研究者によっては,チョッキリゾウムシを独立科として,オトシブミ科と分けて扱うこともある.上部白亜紀から知られるUlyaninaeは,本科の化石群ではないかと考えられている.
キクイムシ科(Scolytidae):口吻は発達しないが,ゾウムシ上科の一群で,樹皮下や材に穿孔することから,森林害虫として有名なものが少なくない.Crowson(1955)は,幼虫がゾウムシ科と区別がつかないとし,キクイムシおよびナガキクイムシをゾウムシ科に含めた.その後,Kuschel(1995)やMarvaldiら(2000, 2002)もこの考えを支持している.一方,Morimoto(1962, 1981)は,頭部構造に根本的な違いが認められることから,ゾウムシ科とは全く別群として扱っている(詳しくは後述する).大半は衰弱木や腐朽木を利用し,ナガキクイムシや一部のキクイムシには,坑道内にアンブロシア菌を栽培し,巣の管理をする養菌性の種も見られる.ナガキクイムシ科(Platypodidae)は通常,独立の科とされるが,Kuschelら(2000)は,ナガキクイムシ科を認めると,キクムシ科が側系統群となるとした.世界から約7,000種が知られ,白亜紀初期の針葉樹の化石から,キクイムシ類の食痕と思われるものが見つかっている.
ミツギリゾウムシ科(Brentidae):近年,本科の定義が徐々に広げられてきており,オーストラリアのEurhynchinae,南アフリカでソテツの種子に産卵するAntliarhininae,サツマイモの大害虫を含むアリモキゾウムシ亜科Cyladinae,ホソクチゾウムシ亜科Apioninae,チビゾウムシ亜科Nanophyinaeに加え,Oberprieler (2000)は幼虫形態からニューヨークゾウムシ亜科Ithycerinaeやアフリカの乾燥地帯に生息し,一見短吻群のゾウムシに似たMicrocerinaeをも本科に含める考えを提唱した.しかし,広義のミツギリゾウムシ科の単系統性はまだ疑問が残り,研究者によってはそれぞれを独立の科とする扱いもある.食性は,狭義のミツギリゾウムシ亜科Brentinaeのように倒木や衰弱木を利用する材食性や腐食性のものから,幼虫が土中で根部を加害するもの(Ithycerinae, Microcerinae, Cyladinae),ゴール形成者や花芽,茎,種子などの植物器官を利用する植食者(Nanophyinae, Apioninae)に至るまで多様である.植食者は,単子葉植物を除く被子植物を主に利用し,一部が針葉樹を利用する.世界から約4,000種が知られ,化石は白亜紀中期以降から見つかっている.
ゾウムシ科(Curculionidae):食性,寄主範囲ともに非常に広く,単子葉植物から双子葉植物にいたる地球上で繁栄した被子植物のあらゆるグループの様々な器官を利用する.また,コケ,シダ,ソテツ,針葉樹など,より原始的な植物をも利用する.狭義のゾウムシ科の分類は混乱しており,100近くの亜科を認める研究者から,10亜科しか認めない研究者まで見解が異なる.近年,ゾウムシ科の中でも,祖先的な交尾器形態をもつ,イボゾウムシ科Brachyceridae,オサゾウムシ科Dryophthoridae(=Rhynchophoridae),イネゾウムシ科Erirhinidaeはそれぞれゾウムシ科から分けて扱われることがある(以下参照).世界から約45,000種が知られる生物界最大の分類単位.化石は白亜紀中期以降から見つかり,第三紀以降多くなる.
イボゾウムシ科:アフリカの乾燥地帯を中心に分布し,太短い口吻と直線状の触角を有し,おもにユリ科などの単子葉植物を利用する.日本産はハスオビコブゾウムシDesmidophorus crassus1種で,アフリカなどに分布する典型的なものとは異なり,別族Desmidophiriniに分類され,アオイ科植物に依存する.
オサゾウムシ科:湿潤な熱帯域を中心に分布し,ヤシ科やショウガ科,ラン科などの単子葉植物を利用する.大形種も少なくない.
イネゾウムシ科:温帯域を中心に分布し,水生,半水性の種を含み,イネ科など水生の単子葉植物を利用する.
系統仮説と高次分類
ゾウムシ上科の分類体系に関する包括的な研究は,Schoenherr(1823-1847)にはじまり,Lacordaire (1863,1866)によって,その基盤が作られた.その後,Crowson(1955)やMorimoto (1962-)らを中心に,修正が加えられてきた.
Kuschel (1995)は分岐学的手法を導入し,ゾウムシ上科内の系統関係を推定し,それに基づく分類体系に関する仮説を提唱した.近年,Marvaldiら(2000, 2002)は,Kuschelの形質データを再分析するとともに,分子データ18SrDNAを導入した解析結果を発表した.ところが,彼女らによる分子データを用いた結果は,ゾウムシ上科内の主要な関係すら明らかにすることができず,最終的には形態データを統合した解析を行い,その影響を大きく受けた解析結果に終わった.
ゾウムシ上科の高次分類や系統関係に関して,いくつかの仮説が提唱されてきているが,その最も大きな相違点は,キクイムシとナガキクイムシの起原に関する見解の違いである.つまり,Crowson(1955)は,両群を幼虫でも区別がつかないとし,ゾウムシ科内に含めた.以後.この見解はLawrence & Newton (1995)をはじめ,Kuschel(1995)やMarvaldiら(2000, 2002)にも指示され,現在,広く普及している(図1, 仮説1).一方,Morimoto(1962,1981)は,頭部の形態構造の明らかな相違から両群をゾウムシ科とは独立の科として扱っている.
前者は,キクイムシ類をゾウムシ上科の中でもっとも高等なグループと考え,生態や外見が類似したゾウムシ科に含まれるキクイゾウムシ亜科から派生したと考えている.ところが,Morimotoは,キクイムシ類とキクイゾウムシ類は,頭部内の構造や口器の構造が基本的に異なり,両群の外見上の類似は,似た生息環境下にあることに起因し,全く別群であるとし,さらにキクイムシ類は,ゾウムシ上科のなかでも比較的初期に分化した一群であると考えている.
Marvaldiらの研究(2000, 2002)は,基本的にKuschel(1995)のデータに基づいた系統解析で,彼等の解析は多数の形態形質に基づいて行われているが,その多くは,分類群間で平行的に生じやすいもので,類縁関係を支持する共有新形質として強固なものは少ない.また,Morimoto (1962, 1981)が指摘した頭部内構造の違いなど,ゾウムシ上科の系統を考える上で,極めて重要と考えられる形質が解析に用いられていないなどの問題点がある.
最近Morimoto & Kojima (2003)は,頭部形態の詳細な観察に基づく系統解析を行った.その結果,頭部形態に以下のような進化傾向が認められた.
1. 扁平な口吻を持ち,大顎や小顎が,水平に動く状態から,高等なゾウムシ類(ミツギリゾウムシ科やゾウムシ科)のように,産卵孔を掘るのに適した円筒状の口吻を獲得し,大顎はやや斜め上方に,小顎はほぼ垂直に動く方向へ変化する.
2. 大顎後関節丘postartisは,大顎に対して斜め下方に向き,短円筒状の状態から,高等ゾウムシに見られるような後関節丘が大顎に対し垂直で,球形をした状態への変化が見られる,これは,postartisが関節する頭部大顎後関節丘postcoilaの状態とも関連し,postcoilaが浅く,前方を向く状態から,高等ゾウムシに見られる球状のpostartisを受ける半球状のくぼみとなり,下口突起hypostomal processも発達する状態への変化傾向が見られる.さらに高等ゾウムシでは,大顎後側面に葉片状の張り出しが有り,外転筋腱基部を覆う.
このような進化傾向が見られる中,キクイムシ類に見られる状態は,祖先的な状態から派生的な状態への移行段階にあり,一般に普及しているキクイムシ類が最も高等なゾウムシであると言う考えは,頭部形態から判断する限り誤りである可能性が高いと考えられた.また,Crowson(1955)により,区別できないとされた幼虫形態に関しても,我々の結果を支持する形質状態が上記研究の過程で見つかった.以上の結果,現段階では,図1に示した仮説2がゾウムシ上科の系統仮説としてもっともらしいものと考えている.なお,ミナミホソクチゾウムシの系統的位置については,現段階で未解決である.
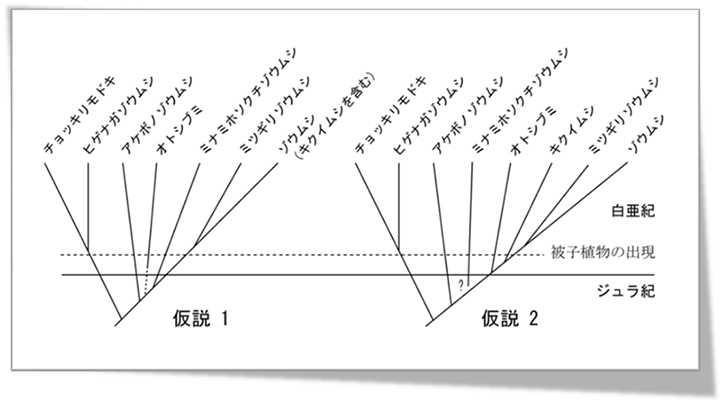
図1.ゾウムシ上科主要科群の系統仮説
系統進化
上記系統仮説に基づき,ゾウムシ上科の系統進化について考察する(図2).
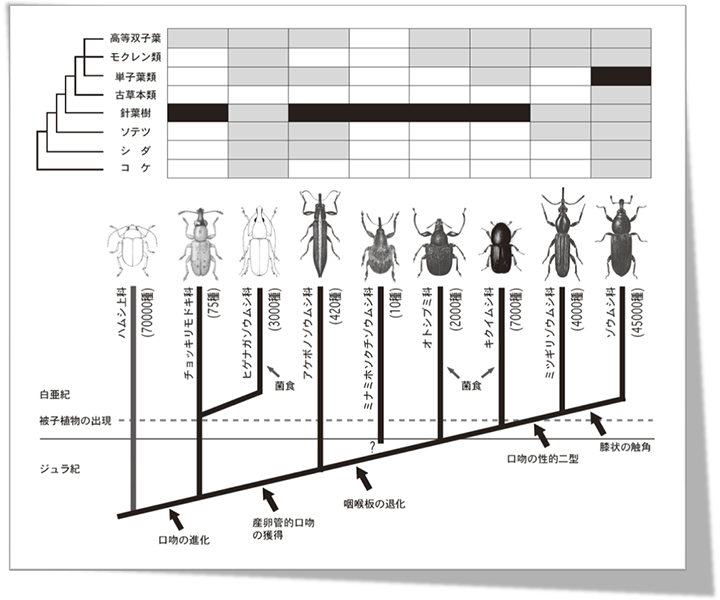
図2.ゾウムシ上科の系統進化と寄主植物との関係
1. 口吻の獲得とゾウムシ上科の分化
確固たるゾウムシ上科の化石として最も古いものは,上部ジュラ紀から得られており,ゾウムシ類の起原は,おそらくジュラ紀中頃にまでさかのぼることができる.その祖先は,当時繁栄していた裸子植物,とくにナンヨウスギなどの針葉樹を利用し,成虫の食性は,花粉食であったと考えられる.この性質は,現存するハムシ類やゾウムシ類で最も祖先的なグループと考えられているミナミナガハムシ亜科Palophaginaeやチョッキリモドキ科に見られる.両群は幼虫もまた,雄球果につき花粉を食べており,寄主植物外部で生活する.そのためチョッキリモドキ科の口吻は,非常に短いものから長いものまで変化に富むが,幼虫が花粉食で,植物外部で生活することから,産卵孔を掘るという目的には用いない.
ヒゲナガゾウムシ科は多くの祖先形質をチョッキリモドキ科と共有するが,幼虫形態からチョッキリモドキ科との姉妹群関係が示され(Marvaldi & Morrone, 2000),産卵管形態も,一部のチョッキリモドキ科と似ている.白亜紀以降に分化し,菌食性を発達させ,熱帯域を中心に多様化した.幼虫は,植物内部に侵入するが,扁平な口吻は,産卵場所に傷を付けるのに用いる程度で,あとは発達した産卵管により卵を産み込む.
2. “産卵管”的口吻の獲得とその進化
アケボノゾウムシ科以降のゾウムシでは,上唇が癒着し,口吻を使って産卵孔を掘る,いわゆる“産卵管”的口吻を獲得する.それに伴い,産卵管は大部分が膜状の構造となる.産卵管的口吻の獲得により,幼虫期間を外敵からの影響が少ない植物器官内部で過ごす生活スタイルを確立した※.
さらに,ミナミホソクチゾウムシ科以降のゾウムシでは,頭部腹面に存在する一対の咽喉縫合線(gular suture)が癒着し,1本の縫合線となる.これに伴い,頭部内で大きな面積を占めていた膜状骨が縮小し,頭部がより球形に近い状態に変化するとともに,大顎や小顎など口器を動かす筋肉が着く,頭部後方の付着面も増大する.
オトシブミ科では,揺籃を作るなど,口吻を複雑な産卵加工に用いるものも出てくる.また,一部には,胞子を貯蔵する器官を備え,菌食性を進化させた.
キクイムシ科およびナガキクイムシ科では,成虫,幼虫ともに材の中で生活するようになり,口吻を発達させなかった.また,アンブロシア菌を貯蔵する器官を備え,養菌性を発達させ多様化した.
一方,ミツギリゾウムシ科やゾウムシ科では,産卵管的口吻を,より円筒形に近い状態に変化させるとともに,雌では,雄に比べ口吻をより細長くする性的二型を発達させ,産卵孔を掘るのにさらに適した頭部形態を獲得した.
さらにゾウムシ科では,これまで直線状であった触角を,1節目を伸長させ膝状に曲がる構造とし,口吻側面の溝に納められる状態とした.これにより,産卵孔を掘る際,触角付着点までしか挿入できない状態から,口吻基部まで挿入可能な口吻を獲得した.
3. 寄主植物との共進化
上記のような産卵管的口吻の獲得とともに,ゾウムシ上科の多様化には,寄主植物との関係が強く影響している.
ジュラ紀から化石記録のあるチョッキリモドキ科,アケボノゾウムシ科,ミナミホソクチゾウムシ科は,その頃繁栄していた裸子植物,とくに針葉樹を利用していたと推定される.その次ぎに分化したと考えられるオトシブミ科も,チョッキリゾウムシの仲間に,少ないながら針葉樹食のものが知られる.キクイムシ科やナガキクイムシ科は,近年の研究から,初期に分化したグループは,針葉樹食であったことが強く示唆されている(Sequeiraら,2000).
一方,ミツギリゾウムシ科では,形態的に古いグループでも,ブナ科(ニューヨークゾウムシ亜科)やアオイ科(Microcerinae)など,被子植物を利用しており,ソテツや針葉樹を利用するものもいるが,本科内ではより高等なグループに含まれる.ゾウムシ科でも,祖先的な交尾器形態を残す分類群は,単子葉植物を利用している.これらのことから,ゾウムシ上科の中での,被子植物食は,ミツギリゾウムシ科以降で起こったと推定される.しかし,ミツギリゾウムシ科の大半は,衰弱木や腐朽木に適応しており,健全な植物組織を利用できるものは,その一部で,完全な植食性が発達するのはゾウムシ科以降である.ミツギリゾウムシ科以降のグループの化石が出てくる年代と,被子植物の出現時期とはほぼ重なる.
Sequeiraら(2000)など,Farrellを中心とする研究グループは,Marvaldiら(2000, 2002)の系統仮説(図1,仮説1)を念頭に,被子植物食を進化させたゾウムシ科の中で,キクイムシ類の針葉樹への適応は,白亜紀以降,被子植物が優勢になるなかで,遺存的に残っている針葉樹を利用することで起こったのではないかと推定している.しかし,我々の系統仮説に基づけば,キクイムシ類までが,元来,針葉樹を寄主としていたと考え,針葉樹食の進化がゾウムシ上科の中で2度独立に起こったことを仮定せずにすみ,より最節約的に説明できる.
ゾウムシ科は,上科全体の約8割を占め,姉妹群であるミツギリゾウムシ科と比べても,10倍以上の種数を有する.ゾウムシ科がこれほどまで多様化した原因は,触角構造の変化を含め,産卵管的口吻の形態や機能を進化させるとともに,単子葉植物から双子葉植物に至る被子植物のあらゆる分類群を利用したことによるものと推測する.
日本のゾウムシ相
日本産のゾウムシ上科は現在,約1,200種(キクイムシ,ナガキクイムシをのぞく)が知られ,日本とほぼ同じ面積で,研究の進んでいるイギリスの2倍,中部ヨーロッパ全域に分布するゾウムシの種数におよぶ.最大の分類群であるゾウムシ科は約850種を含み,チョウ類の2.5倍,分類の進んだカミキリムシ科にほぼ匹敵する数が,すでに知られていることになる.しかし,今後,500種以上の追加が見込まれていて,最終的には,2,000種近くになると予想される.ゾウムシ類の繁栄の要因は,産卵管的機能を有した口吻の獲得と同時に,様々な状態の,植物官ごとに分化した幅広い植物資源の利用が挙げられる.
植物と密接に関係しながら進化してきたグループで,そのファウナを捉える際,植生が重要な要因となる.日本産ゾウムシ相の特徴として,草本性植物より,木本性植物に依存した種が多いという傾向が見られる.これは,日本では草原環境に比べ,森林環境が発達していることに起因する.木本性植物に依存した種には,材食性,腐材食性のヒゲナガゾウムシ科,クチカクシゾウムシ亜科,アナアキゾウムシ亜科,キクイゾウムシ亜科や,生木につく,ノミゾウムシ族,シギゾウムシ族などのゾウムシ亜科が挙げられる.これらの分類群は,国内で多様な種に分化していて,日本産だけでも,ヨーロッパ全体の種数よりはるかに多い.一方,草本性の種を多く含むホソクチゾウムシ科や,サルゾウムシ族,コガタゾウムシ族,タコゾウムシ族では,種多様性が低い.
参考文献
- - Crowson, R. A., 1955. The natural classification of the families of Coleoptera. Lloyd and Co., London.
- - Lacordaire, J. T., 1863, 1866. Historie naturelle der insects. Genera des Coleopteres …, vol. 6 (1863), 7 (1866).
- - Lawrence, J. F. & A. F., Newton, 1995. In. Pakaluk & Slipinski (Ed.): Biology, phylogeny, and classification of Coleoptera, Vol. 2: 778-1006.
- - Schoenherr, C. J., 1823. Isis, Jena , 7: 1132-1146.
- - Schoenherr, C. J., 1826. Curculionidum dispositio methodica …, Pt. 4. Fleischer, Lipsiae.
- - Schoenherr, C. J., 1833-1845. Genera et species curculionidum. Vol. 1-8.
- - Marvaldi, A.E. & J.J. Morrone, 2000. Insect Syst. Evol., 31: 43-58.
- - Marvaldi, A.E. et al, 2002. Syst. Biol., 51: 761-785.
- - 小島弘昭,2004.昆虫と自然,39(4):22-26.
- - 小島弘昭,2008.月刊むし,(443): 17-27.
- - Kuschel, G., 1995. Mem. Entomol. Soc. Wash., 14: 5-33.
- - Kuschel, G. et al., 2000. Invert. Taxon., 14(6): 771-805.
- - Morimoto, K., 1962. J. Fac. Agric. Kyushu Univ., 12: 21-66.
- - Morimoto, K., 1981. Akitu (N. S.), 34: 1-10.
- - Morimoto, K. & H. Kojima, 2003. Esakia, (43): 133-169.
- - Morimoto, K. et al., 2006. The Insects of Japan, 3. 406 pp. Touka Shobo, Fukuoka.
- - Oberprieler, R., 2000. Invert. Taxon., 14(6): 755-770.
- - Sequeira, A.S. et al., 2000. Proc. R. Entomol. Soc. Lond., B 267: 2359-2366.
※ ゾウムシ科やミツギリゾウムシ科では,二次的に口吻を短くさせ,産卵の際,口吻を用いないものや,幼虫が植物外部で生活するものも一部に出てくる.